
Peptoids: Emerging Therapeutics for Neurodegeneration
Lauren M. Wolf,1 Shannon L. Servoss,2 and Melissa A. Moss1,3,*
Abstract
In recent decades, the increasing prevalence of age-associated neurodegenerative diseases has underscored the need for targeted therapeutic strategies and novel diagnostics. Peptide-based neurotherapeutics offer high specificity and tolerability but are limited by proteolytic degradation in vivo. Peptoids, or N-substituted glycines, are versatile peptidomimetics that evade proteolytic degradation yet maintain many qualities that render peptides attractive neurotherapeutic candidates. These molecules may be engineered to their application through modifications that enhance structural stability and reactivity and can withstand various physiological stressors to retain their intended function within anomalous microenvironments.
Peptoids generally demonstrate greater cellular permeability than their corresponding peptides, are less immunogenic, and can be administered intranasally, all properties that enhance their potential as neurotherapeutics. Peptoids have primarily been explored as aggregation inhibitors to prevent the deleterious protein plaque deposition associated with several neurodegenerative disorders. However, novel research has uncovered the potential of peptoids toward additional neurotherapeutic applications. Peptoids can modulate cell signaling pathways involved in axonal function and current modulation and can block cell signaling events associated with apoptosis. In addition, these peptidomimetics are able to function as anti- inflammatory agents via multiple mechanisms. Moreover, the versatility and low cost of peptoids render them ideal instruments in biomarker detection, discovery, and imaging. This mini-review explores these diverse applications of peptoids within the context of neurodegenerative disease.
Introduction
The increasing prevalence of neurodegenerative diseases that accompany an aging population, such as Alzheimer’s disease (AD), Huntington’s disease (HD), and Parkinson’s disease (PD), underscores a need for effective therapeutic strategies. An improved understanding of the functional relevance of protein-protein interactions in homeostasis and disease pathologies has boosted the value of peptides as neurotherapeutic agents1–3. Peptides can be synthesized to achieve binding specificity similar to native proteins, making them favorable for therapeutic strategies that disrupt or replace protein-protein interactions as well as broader applications, such as subcellular targeting strategies and detection of biomarkers for central nervous system (CNS) pathologies4,5. Nonetheless, the practical application of peptide therapeutics in vivo is limited by vulnerability to proteases. While structural alterations can be introduced to increase peptide half-life, including D-amino acid substitution6, terminal acetylation or amidation7, and modification of the proteolytic site8, these alterations often reduce therapeutic activity6–8.
Peptoids, or oligomers of N-substituted glycines, are peptidomimetics that evade proteolytic degradation via repositioning of the side-chain from the α-carbon to the amide nitrogen (Figure 1)9. Peptoids are synthesized with relative ease and at comparatively low cost10,11. While peptoids can be constructed to mimic peptides in side-chain chemistry, a direct sequence translation is not always the most effective approach. Instead, the versatility of peptoids is often exploited through design, such as the incorporation of reactive sites or the conferment of structural stability through inclusion of chiral, aromatic side chains or cyclization of the peptoid9,12–15. Peptoids exhibit increased cellular permeability as compared to the corresponding peptide, thus enhancing the potential for intracellular targets. Intranasal administration of peptoids also achieves effective delivery to the CNS, enabling use as a therapeutic for neurodegenerative diseases16–18. Additionally, peptoids are less likely than peptides to elicit an immunogenic response while retaining the ability to specifically target and inhibit protein-protein interactions19,20. Together, these characteristics render peptoids an attractive, novel neurotherapeutic agent.
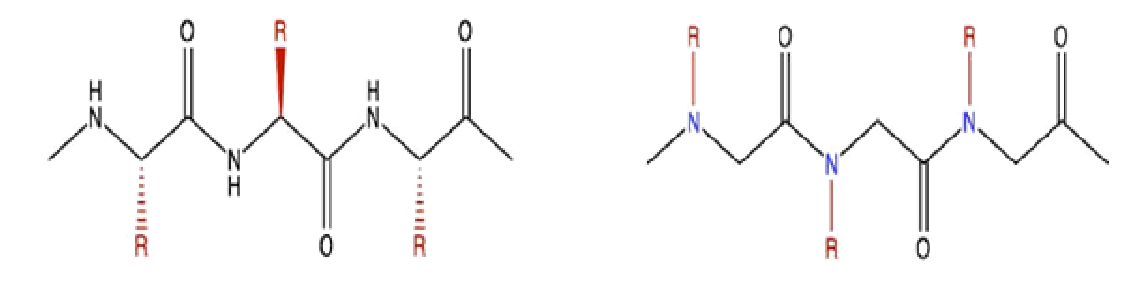
Figure 1. Structural differences between peptides and peptoids.Although their backbones are similar, side chains are attached to the α-carbon in peptides (panel a), while peptoids have side chains attached to the amide nitrogen (panel b). This modification confers resistance to proteolytic degradation.
Peptoids as Aggregation Inhibitors
Several neurodegenerative diseases are amyloidoses, wherein a harmless protein monomer self-assembles to form toxic aggregates that possess a highly-ordered cross-β structurei21-23. AD, HD, and PD are neurodegenerative amyloidoses associated with aggregation of amyloid-β (Aβ), Huntingtin, and α-synuclein proteins, respectively. Pancreatic amylin amyloid is a common disorder in Type II diabetes; however, increased amylin amyloid has also been detected in the cerebrovasculature of late-onset AD patients who lack diagnoses for clinical diabetes21,23. One therapeutic strategy for neurodegenerative amyloidoses is inhibition of aggregation or aggregate destabilization. Several peptoids have been explored as therapeutic agents toward this approach.
Starting from an on-bead library of more than 38,000 unique peptoid sequences containing four variable residues, Luo et al. identified a peptoid selective for Aβ1-42, the more aggregation-prone form of the AD-associated amyloidogenic protein. Both the monomer and dimer of this peptoid inhibit Aβ1-42 aggregation in a concentration-dependent manner. Moreover, the dimer demonstrates high affinity for Aβ1-42 and also confers neuroprotection in an amyloid toxicity assay24. A distinct anti-amyloidogenic peptoid also exhibiting high affinity for Aβ1-42 was identified via surface plasmon resonance from a library of more than 4,000 six-mer peptoids containing four variable residues. This peptoid inhibits both early stage oligomerization as well as the formation of mature aggregates and protects against Aβ-induced toxicity25. Through rational design, an eight-mer peptoid was developed to both mimic the five-residue hydrophobic core of Aβ and incorporate additional aromatic residues. This peptoid significantly attenuates the formation of fibrillar Aβ1-40, the most abundant form of the AD-associated amyloidogenic protein24. A subsequent study used variants of this peptoid to examine the importance of side-chain placement and side-chain chirality on inhibition of Aβ1-40 aggregation. Although both variants significantly reduce Aβ1-40 aggregation, the absence of side-chain chirality leads to changes in the size and morphology of the aggregates formed, which have the potential to modulate aggregate toxicity13,26. Two peptide-peptoid hybrids, designed to both mimic the Aβ hydrophobic core and incorporate N-terminal hydrogen bond donors, also inhibit aggregation of Aβ1−42 in a manner superior to their peptide counterparts. These peptide-peptoid hybrids additionally demonstrate stability in cell culture and attenuate Aβ toxicity27.
While inhibition of Aβ aggregation by peptoids has been the most widely studied among neurodegenerative amyloidogenic proteins, peptoid inhibitors have also been explored within other amyloid-associated disease models. Chen et al. identified a peptoid that significantly inhibits aggregation of mutant polyglutamine -expanded Huntingtin fragments (mHtt). Intracerebroventricular infusion of the peptoid for 30 days reduced mHtt aggregate formation, stabilized Ca2+ signaling, and reduced neuronal apoptosis in a mouse model of HD28. In a study targeting amyloid formation by amylin, a rationally-designed peptoid was developed to mimic amylin residues 20-29. This peptoid inhibits aggregation of the native amylin fragment in vitro29. Overall, it is interesting to note that each of the peptoids reported to inhibit amyloid protein aggregation contains aromatic side chains, suggesting the importance of this structural element within the recognition and disruption processes.
Peptoids as Effectors of Cell Signaling
The intervention and modulation of cell signaling pathways offers an additional approach to CNS therapeutic development through disruption of deleterious signaling events. Protein-protein interactions that drive these pathways are potential therapeutic targets, and peptoids as well as peptide-peptoid hybrids have been explored as effectors of these interactions4,30. A peptoid capable of modulating semaphorin 3A (Sema3A), a negative regulator of axon guidance that is overexpressed following CNS injury, was developed to promote axonal regeneration and reduce neural scarring. By blocking the interaction between glycosaminoglycans and Sema3A to attenuate Sema3A activity, this peptoid enhances neuronal regrowth in vitro31–33. Therapeutic inhibition of histone deacetylase-6 (HDAC6) presents promise for neurodegenerative diseases, as evidenced by observations that HDAC6 inhibition rescues impaired axonal transport and HDAC6 knockdown improves function in a mouse model of AD34,35. A recent study used rational design to identify a peptoid-based inhibitor of HDAC6. Although not yet investigated in neurodegeneration, this inhibitor is an effective chemosensitizing agent, a function for which HDAC6 has also emerged as a therapeutic target36. Peptide-peptoid hybrids were investigated as potential antagonists for neurotensin receptor 2, a key receptor in the regulation of tonic pain sensitivity and some psychiatric diseases. The hybrids demonstrate a higher affinity for the receptor and inhibit constitutive mitogen-activated protein kinase (MAPK) activity stronger than the natural ligand neurotensin37. Peptide-peptoid hybrid analogs of KIIIA, an endogenous voltage-gated sodium channel blocker, were developed to maintain high binding affinity but reduce the current blocked, thus providing a greater margin of safety for use as an analgesic. An N-methylglycine point substitution successfully lowers the blockage of current by 20%, creating a peptide-peptoid hybrid with more desirable functionality than naturally occurring KIIIA38. Although direct applications to neurodegeneration remain to be extensively explored, peptoids are well-established as effective modulators of cell signaling within the CNS thus exemplifying their potential in this capacity toward therapies involving neurodegenerative diseases.
Peptoids as Neuroprotective Agents
By targeting apoptotic signaling pathways, peptoids may also protect against neuronal loss. Two trialkylglycine peptoids confer neuroprotection against excitotoxicity-induced apoptosis, a common element in neurodegenerative pathogenesis. This protection correlates with diminished activation of caspase-3 without compromising N-methyl D-aspartate (NMDA) receptor activity, thus circumventing deleterious side effects of current drugs39. Peptoids have also been used as antagonists for apoptotic protease-activating factor-1 (Apaf-1) to render the apoptosome complex inactive40,41. Such antagonism impairs mitochondrial-mediated apoptosis, which is associated with many neurodegenerative diseases42. Introducing conformational constraint43 and conjugation of the peptoid to cell penetrating peptides44 or a polymeric carrier45 enhance the effectiveness of these peptoids. Thus, peptoids can target neuroprotective pathways relevant to neurodegenerative diseases.
Peptoids for Attenuation of Neuroinflammation
Inflammation plays an active role in the development and progression of neurodegeneration, and suppression of neuroinflammation has become a target for therapeutic development46,47. Peptoids have been proposed as an avenue for manipulation of antigen-specific immune responses by functioning as “antigen surrogates” to amplify a weak immune response or weaken a deleterious immune response through competition. Peptoids that incorporate backbone chirality to restrict conformation have been selected for high binding affinity and selectivity to the antigen-binding site of therapeutically relevant antibodies, demonstrating the feasibility of this approach48. In a different approach, peptoid mimics of catalase and superoxide dismutase have been designed to mitigate the impact of oxidative stress, an integral component of inflammation. These peptoids bind redox-active metal ions to form complexes capable of scavenging and detoxifying reactive oxygen and nitrogen species49,50. To modulate activation of immune cells, lipidated peptide-peptoid hybrids have been identified as both agonists51 and antagonists52,53 of formyl peptide receptor 2 (FRP2), expressed by neutrophils and involved in neutrophil activation. Upon binding FRP2, antagonists are able to attenuate FRP2-specific activation of neutrophils52,53. These examples demonstrate that peptoids hold promise for attenuating neurodegeneration-associated inflammation via a variety of mechanisms.
Peptoids for Biomarker Detection and Imaging
The symptomatic and pathological overlap across many neurodegenerative diseases complicates their diagnosis, and the inaccessibility and complexity of the CNS present challenges for diagnostic assays54. When coupled with the difficulty of reversing neuronal damage, which implicates early intervention as a key to successful therapeutics, these circumstances underscore the need for novel biomarker discovery and diagnostic tools. Through their specificity and stability, relatively low cost, and ease of modification, peptoids may function as biomimetic receptors with diagnostic applications4,5,12,55.
A combinatorial library of peptoids can be readily created to provide a random, diverse set of molecules from which to select a capture agent for biomarkers within an accessible biological fluid. This approach has been used to discover a peptoid ligand with nanomolar binding affinity that is selective for Aβ1-42 (vs. Aβ1-40), a suggested biomarker for AD24. By applying a peptoid library to compare antibody capture between patient and control samples, an antibody-peptoid ligand set can also be identified without prior knowledge of the antibody biomarker itself. Such a strategy offers a distinct advantage for many neurodegenerative diseases wherein effective biomarkers have not been identified and antibodies are suggested as prospective biomarker candidates56,57. In this manner, potential IgG antibody biomarkers have been discovered within an animal model of multiple sclerosis and human subjects with AD58 and PD59. Peptoids may also be leveraged for imaging applications to add specificity to contrast and imaging agents. A potent peptoid antagonist for vascular endothelial growth factor (VEGF) receptor 2 was labeled with a positron emitter for successful positron emission tomography (PET) imaging of the receptor in vivo60. While Hao et al. applied this technology for tumor imaging, parallel approaches could be developed using peptoid ligands for neurodegeneration biomarkers, which may range from altered receptor expression to amyloid deposition.
Conclusion and Future Perspectives
Peptoids were first proposed in 1992 with the original intent of advancing the drug discovery process9. These molecules offer a great deal of potential as therapeutics for neurodegeneration and for applications beyond this sphere. Already, peptoids have been explored as agents for anti-aggregation, neuroprotection, anti-inflammation, biomarker detection and discovery, and imaging, as summarized in Table 1. Their stability, specificity, and enhanced permeability equip peptoids with the therapeutic advantages of both small molecules and peptides. Subcellular targeting facilitates treatment, and structural stability allows peptoid functionality to be retained in extreme physiological environments. For example, β-secretase, the rate-limiting enzyme in Ab release from its protein precursor, is activated within the acidic climate of early endosomes; peptoids exemplify β-secretase inhibitor candidates with the potential for both targeted delivery to this compartment and stability within this environment4. Their size and capacity for intranasal delivery also make peptoids a compelling prospect for neurological interventions16–18. In addition to their many current potential applications, the possibilities for these degradation-resistant protein-mimetics grow with increased knowledge of peptoid assembly and folding behavior.
| Binding Target | Intervention | Source(s) |
|---|---|---|
| Amylin | Inhibition of aggregation | 29 |
| Aβ1-40 | Inhibition of aggregation | 13 |
| Aβ1-42 | Inhibition of aggregation; Inhibition of Aβ neurotoxicity | 24,25 |
| Aβ1-42 | peptide-peptoid hybrid; Inhibition of aggregation; Inhibition of Aβ neurotoxicity | 27 |
| Apaf-1 | Neuroprotection via inhibition of mitochondrial-mediated apoptosis | 40,41,44,45 |
| Apaf-1 | inhibition of apoptosome formation | 43 |
| oxidative stress by-products | catalase and superoxide dismutase mimic; neutralization of reactive oxygen and nitrogen spices | 49,50 |
| FRP2 | peptide-peptoid hybrid; Receptor antogonist; attenuation of neutrophil activation | 51-53 |
| HDAC6 | Inhibition of HDAC6 enzymatic activity | 36 |
| mHtt | Inhibition of aggregation; protection against neutroxicity; stabilization of ca2+ signaling | 28 |
| IgG antibodies | Biomarker detection | 58,59 |
| Voltage-gated sodium channels | peptide-peptoid hybrid; Blockage of current | 38 |
| NTS2 | peptide-peptoid hybrid; Receptor antogonist; Inhibition of MAPK enzymatic activity | 37 |
| Sema3A | Promotion of axonal regrowth | 31,33 |
| Unknown | Neuroprotection correlated with reduced caspase 3 activity | 39 |
| VEGF receptor 2 | in vivo imaging | 60 |
Table 1Prospective neurotherapeutic applications of peptoids and peptide-peptoid hybrids.
Conflict of Interest
The authors report no competing interests.
References
- Alavijeh MS, Chishty M, Qaiser MZ, et al. Drug metabolism and pharmacokinetics, the blood-brain barrier, and central nervous system drug discovery. J Am Soc Exp Neurother. 2005; 2(4): 554-571.
- Wójcik P, Berlicki L. Peptide-based inhibitors of protein-protein interactions. Bioorganic Med Chem Lett. 2016; 26(3): 707-713.
- Kumar J, Namsechi R, Sim VL. Structure-based peptide design to modulate amyloid beta aggregation and reduce cytotoxicity. PLoS One. 2015; 10(6): 1-18.
- Rajendran L, Knölker HJ, Simons K. Subcellular targeting strategies for drug design and delivery. Nat Rev Drug Discov. 2010; 9(1): 29-42.
- Scarano S, Lisi S, Ravelet C, et al. Detecting Alzheimer’s disease biomarkers From antibodies to new bio-mimetic receptors and their application to established and emerging bioanalytical platforms - A critical review. Anal Chim Acta. 2016; 940: 21-37.
- Hong SY, Oh JE, Lee KH. Effect of D-amino acid substitution on the "stability," the secondary "structure," and the activity of membrane-active peptide. Biochem Pharmacol. 1999; 58(99): 1775-1780.
- John H, Maronde E, Forssmann WG, et al. N-terminal acetylation protects glucagon-like peptide GLP-1-(7-34)-amide from DPP-IV-mediated degradation retaining cAMP- and insulin-releasing capacity. Eur J Med Res. 2008; 13(2): 73-78.
- Strömstedt AA, Pasupuleti M, Schmidtchen A, et al. Evaluation of strategies for improving proteolytic resistance of antimicrobial peptides by using variants of EFK17, an internal segment of LL-37. Antimicrob Agents Chemother. 2009; 53(2): 593-602.
- Zuckermann RN, Kerr JM, Kent SB, et al. Efficient method for the preparation of peptoids "[oligo (N-substituted glycines)]" by submonomer solid-phase synthesis. J Am Chem Soc. 1992; 114(26): 10646-10647.
- Fowler SA, Blackwell HE. Structure-function relationships in peptoids: recent advances toward deciphering the structural requirements for biological function. Org Biomol Chem. 2009; 7(8): 1508-1524.
- Horn T, Lee BC, Dill KA, et al. Incorporation of chemoselective functionalities into peptoids via solid-phase submonomer synthesis. Bioconjug Chem. 2004; 15(2): 428-435.
- Zuckermann RN, Kodadek T. Peptoids as potential therapeutics. Curr Opin Mol Ther. 2009; 11(3): 299-307.
- Turner JP, Chastain SE, Park D, et al. Modulating amyloid-β aggregation: The effects of peptoid side chain placement and chirality. Bioorg Med Chem. 2017; 1(1): 1-7.
- Gorske BC, Blackwell HE. Tuning peptoid secondary structure with pentafluoroaromatic functionality: A new design paradigm for the construction of discretely folded peptoid structures. J Am Chem Soc. 2006; 128(44): 14378-14387.
- Shin SBY, Yoo B, Todaro LJ, et al. Cyclic peptoids. J Am Chem Soc. 2007; 129(11): 3218-3225.
- Ross TM, Zuckermann RN, Reinhard C, et al. Intranasal administration delivers peptoids to the rat central nervous system. Neurosci Lett. 2008; 439(1): 30-33.
- Tan J, Cheong H, Park YS, et al. Cell-penetrating peptide-mediated topical delivery of biomacromolecular drugs. Curr Pharm Biotechnol. 2014; 15(1): 231-239.
- Bentz J, Hill B, Illum L. Propylene glycol compositions for intranasal administration of active agents to the central nervous system. U.S. Patent Application 20150010557 A1, issued January 8, 2015.
- Kwon YU, Kodadek T. Quantitative evaluation of the relative cell permeability of peptoids and peptides. J Am Chem Soc. 2007; 129(6): 1508-1509.
- Patch JA, Kirshenbaum K, Seurynck SL, et al. Versatile oligo(N-substituted) glycines: The many roles of peptoids in drug discovery. In: Pseudo-Peptides in Drug Development. Wiley-VCH Verlag GmbH & Co. KGaA. 2005; 1-31.
- 2Jackson K, Barisone GA, Diaz E, et al. Amylin deposition in the brain: A second amyloid in Alzheimer disease. Ann Neurol. 2013; 74(4): 517-526.
- 2Francioso A, Punzi P, Boffi A, et al. β-Sheet interfering molecules acting against β-amyloid aggregation and fibrillogenesis. Bioorganic Med Chem. 2015; 23(8): 1671-1683.
- Knowles TPJ, Vendruscolo M, Dobson CM. The amyloid state and its association with protein misfolding diseases. Nat Rev Mol Cell Biol. 2014; 15(6): 384-396.
- Luo Y, Vali S, Sun S, et al. Aβ42-binding peptoids as amyloid aggregation inhibitors and detection ligands. ACS Chem Neurosci. 2013; 4(6): 952-962.
- Zhao Z, Zhu L, Li H, et al. Antiamyloidogenic activity of Aβ42-binding peptoid in modulating amyloid oligomerization. Small. 2017; 13(1): 1602857.
- Servoss S, Moss M. Peptoids and methods for treating Alzheimer’s disease. U.S. Patent 8,809,275, issued August 19, 2014.
- Rajasekhar K, Suresh SN, Manjithaya R, et al. Rationally designed peptidomimetic modulators of Aβ toxicity in Alzheimer’s disease. Sci Rep. 2015; 5: 8139.
- Chen X, Wu J, Luo Y, et al. Expanded polyglutamine-binding peptoid as a novel therapeutic agent for treatment of Huntington’s disease. Chem Biol. 2011; 18(9): 1113-1125.
- Elgersma RC, Mulder GE, Kruijtzer JAW, et al. Transformation of the amyloidogenic peptide amylin(20-29) into its corresponding peptoid and retropeptoid: Access to both an amyloid inhibitor and template for self-assembled supramolecular tapes. Bioorganic Med Chem Lett. 2007; 17(7): 1837-1842.
- Stone TA, Deber CM. Therapeutic design of peptide modulators of protein-protein interactions in membranes. Biochim Biophys Acta - Biomembr. 2016; 1859(4): 577-585.
- Corredor M, Bonet R, Moure A, et al. Cationic peptides and peptidomimetics bind glycosaminoglycans as potential Sema3A pathway inhibitors. Biophys J. 2016; 110(6): 1291-1303.
- Mecollari V, Nieuwenhuis B, Verhaagen J. A perspective on the role of class III semaphorin signaling in central nervous system trauma. Front Cell Neurosci. 2014; 8(October): 328.
- Montolio M, Messeguer J, Masip I, Guijarro P, Gavin R, del Río JA, Messeguer A, Soriano E. A semaphorin 3A inhibitor blocks axonal chemorepulsion and enhances axon regeneration. Chem Biol. 2009;16(7):691-701.
- Kim JY, Woo SY, Hong YB, et al. HDAC6 inhibitors rescued the defective axonal mitochondrial movement in motor neurons derived from the induced pluripotent stem cells of peripheral neuropathy patients with HSPB1 mutation. Stem Cells Int. 2016; 2016: 1-14.
- Simoes Pires C, Zwick V, Nurisso A, et al. HDAC6 as a target for neurodegenerative diseases: What makes it different from the other "HDACs?" Mol Neurodegener. 2013; 8: 7.
- Diedrich D, Hamacher A, Gertzen CGW, et al. Rational design and diversity-oriented synthesis of peptoid-based selective HDAC6 inhibitors. Chem Commun. 2016; 52(15): 3219-3222.
- Einsiedel J, Held C, Hervet M, et al. Discovery of highly potent and neurotensin receptor 2 selective neurotensin mimetics. J Med Chem. 2011; 54(8): 2915-2923.
- Walewska A, Han TS, Zhang MM, et al. Expanding chemical diversity of conotoxins Peptoid-peptide chimeras of the sodium channel blocker µ-KIIIA and its selenopeptide analogues. Eur J Med Chem. 2013; 65: 144-150.
- Montoliu C, Humet M, Canales JJ, et al. Prevention of in vivo excitotoxicity by a family of trialkylglycines, a novel class of neuroprotectants. J Pharmacol Exp Ther. 2002; 301(1): 29-36.
- Malet G, Martín AG, Orzáez M, et al. Small molecule inhibitors of Apaf-1-related caspase-3/-9 activation that control mitochondrial-dependent apoptosis. Cell Death Differ. 2006; 13(9): 1523-1532.
- Mondragón L, Orzáez M, Gortat A, et al. Molecules that bind a central protein component of the apoptosome Apaf-1 and modulate its activity. In: Cecconi F, D’Amelio M, eds. Apoptosome An Up-and-Coming Therapeutical Tool. Dordrecht Springer Netherlands. 2010; 75-94.
- Arun S, Liu L, Donmez G. Mitochondrial biology and neurological diseases. Curr Neuropharmacol. 2016; 14(2): 143-154.
- Corredor M, Bujons J, Orzáez M, et al. Optimizing the control of apoptosis by amide/triazole isosteric substitution in a constrained peptoid. Eur J Med Chem. 2013; 63: 892-896.
- Orzáez M, Mondragón L, Marzo I, et al. Conjugation of a novel Apaf-1 inhibitor to peptide-based cell-membrane transporters: Effective methods to improve inhibition of mitochondria-mediated apoptosis. Peptides. 2007; 28(5): 958-968.
- Vicent MJ. Polymer-drug conjugates as modulators of cellular apoptosis. AAPS J. 2007; 9(2): E200-E207.
- Salminen A, Huuskonen J, Ojala J, et al. Activation of innate immunity system during aging: NF-κB signaling is the molecular culprit of inflamm-aging. Ageing Res Rev. 2008; 7(2): 83-105.
- Kempuraj D, Thangavel R, Natteru PA, et al. Neuroinflammation induces neurodegeneration. J Neurol Neurosurg Spine. 2016; 1(1): 1003.
- Kodadek T. Chemical tools to monitor and manipulate the adaptive immune system. Chem Biol. 2014; 21(9): 1066-1074.
- Fisher AEO, Maxwell SC, Naughton DP. Catalase and superoxide dismutase mimics for the treatment of inflammatory diseases. Inorg Chem Commun. 2003; 6(9): 1205-1208.
- Fisher AEO, Naughton DP. Novel peptoids for the detection and suppression of reactive oxygen and nitrogen species. Biochem Soc Trans. 2003; 31(Pt 6): 1302-1304.
- Holdfeldt A, Skovbakke SL, Winther M, et al. The lipidated peptidomimetic Lau-((S)-Aoc)-(Lys-βNphe)6-NH2 is a novel formyl peptide receptor 2 agonist that activates both human and mouse neutrophil NADPH oxidase. J Biol Chem. 2016; 291(38): 19888-19899.
- Skovbakke SL, Heegaard PMH, Larsen CJ, et al. The proteolytically stable peptidomimetic Pam-(Lys-βNSpe)6-NH2 selectively inhibits human neutrophil activation via formyl peptide receptor 2. Biochem Pharmacol. 2015; 93(2): 182-195.
- Skovbakke SL, Winther M, Gabl M, et al. The peptidomimetic Lau-(Lys-βNSpe)6-NH2 antagonizes formyl peptide receptor 2 expressed in mouse neutrophils. Biochem Pharmacol. 2016; 119: 56-65.
- Jellinger KA. Cerebral multimorbidity in aging. J Neurol Neuromedicine. 2016; 1: 1-5.
- Vanderstichele H, Kodadek T. Roadblocks for integration of novel biomarker concepts into clinical routine The peptoid approach. Alzheimers Res Ther. 2014; 6(2): 23.
- Nagele E, Han M, DeMarshall C, et al. Diagnosis of Alzheimer’s disease based on disease-specific autoantibody profiles in human sera. PLoS One. 2011; 6(8): 1-7.
- Han M, Nagele E, DeMarshall C, et al. Diagnosis of Parkinson’s disease based on disease-specific autoantibody profiles in human sera. PLoS One. 2012; 7(2): e32383.
- Reddy MM, Wilson R, Wilson J, et al. Identification of candidate IgG biomarkers for Alzheimer’s disease via combinatorial library screening. Cell. 2011; 144(1): 132-142.
- Yazdani U, Zaman S, Hynan LS, et al. Blood biomarker for Parkinson disease peptoids. Npj Park Dis. 2016; 2: 16012
- Hao G, Hajibeigi A, León Rodríguez LM De, et al. Peptoid-based PET imaging of vascular endothelial growth factor receptor (VEGFR) expression.