
Human Gastrokine 1 and its anti-amyloidogenic properties
Chiara Stella Di Stadio1, Filomena Altieri1, Giuseppina Minopoli1, Giuseppina Miselli1, Emilia Rippa1*, Paolo Arcari1,2*
Abstract
Gastrokine 1 (GKN1) is a 18 kDa stomach protein highly expressed in normal gastric tissue but absent in gastric cancer. GKN1 plays its major role in maintaining gastric mucosal integrity. Because of the presence in its central region of a BRICHOS domain, GKN1 is characterized by multifunctional properties since it interacts and regulates the activity of several proteins. The BRICHOS domain consists of about 100 amino acids and has been found in protein families often associated with major human diseases like familial British and Danish dementia (BRI2) or respiratory distress syndrome (surfactant protein C) (SP-C), both associated with amyloid formation. It has been shown that BRICHOS is a chaperon domain that has the property of binding precursor protein regions with high β-sheet tendencies, thereby preventing them from amyloid formation. Like the BRICHOS domains from BRI2 and SP-C precursor (proSP-C), also GKN1 is able to prevent fibrils formation of amyloid-beta peptide (Aβ) and to interact with the C-terminal region of APP thus hindering the γ-secretase proteolytic sites. Indeed, amyloid is of great medical importance since it originates in several major fatal diseases such as Alzheimer, Parkinson and diabetes mellitus. The results collected until now on the BRICHOS properties of GKN1 and those from other BRICHOS suggest that the different amyloids recognized by BRICHOS should contain similar structural elements therefore, the BRICHOS domain represents a potential powerfull tool for therapeutic approaches against amyloid associated diseases.
Abbreviations: AD, Alzheimer disease; Aβ, amyloid-beta peptide; APP, amyloid precursor protein; CTFs, APP C-Terminal Fragments; GC, gastric cancer; DAPT, N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester; DMSO, dimethyl sulfoxyde; GC, gastric cancer; GKN1, gastrokine 1; flGKN1, full-lenght GKN1; MS, mass spectrometry; rGKN1, recombinant GKN1; SPR, surface plasma resonance; TM, transmembrane; TMpred, transmembrane prediction.
Introduction
Gastrokine1 (GKN1), a 18 kDa protein also known as antrum mucosal protein (AMP-18)1, is highly expressed in the gastric mucosa of many mammals species1,2. The gene coding GKN1 (CA11), located on the chromosome 2p13 (accession number: BK0017373), is about 6 kb long and contains 6 exons. The nucleotide sequence of GKN1 shows the presence of two translation starting sites (ATG) however, only from the second ATG the gene would encode for a protein of 185 amino acids. Edman’s degradation performed on native human GKN1 confirmed the assumption and showed that the protein contains in its N-terminal region a 20 amino acids extracellular signal peptide3 (Fig. 1).
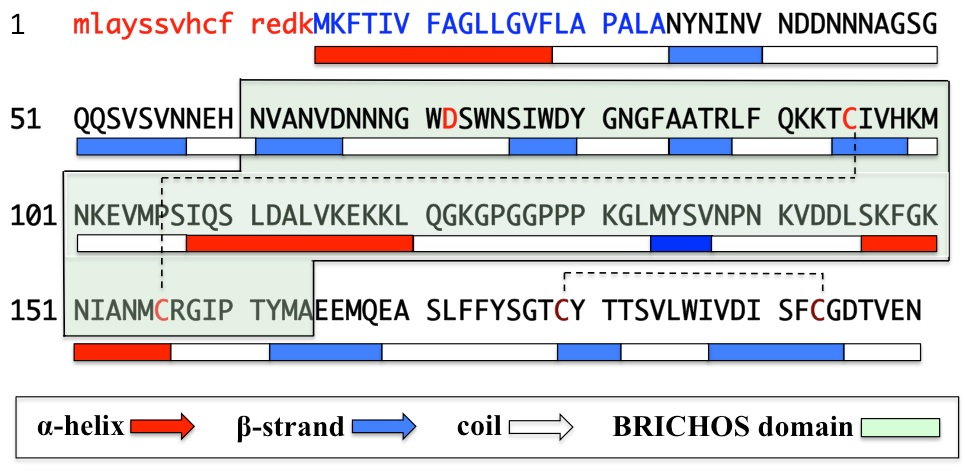
Fig. 1. Amino acid sequence and structure of human GKN1.Peptide highlighted in red (lower case) is that generated from the first ATG but is not translated. Sequence highlighted in blue is the 20 amino acids leader peptide. Sequence highlighted in black is the mature GKN1. Conserved D72, and BRICHOS domain C95 and C156 are highlighted in red. Conserved C179 and C193 are highlighted in brown. The predicted secondary structure of GKN1 was obtained at PSIPRED Protein Structure Prediction Server. Dashed lines represent the disulfide bridges.
GKN1 plays a central role in maintaining the integrity of the gastric mucosa and to facilitate its repair after damage4,5. The protein is localized mainly within the granules just under the apical plasma membrane. This finding suggests that GKN1 is primarly a secreted protein. It has been estabilished that the GKN1 mRNA is abundant only in normal human stomach and in particular in cardia, body and antrum, but absent or almost undetectable in gastric adenocarcinomas; gastro-oesophageal adenocarcinoma cell line and other normal and tumor gastro-intestinal tissues6. Therefore, GKN1 gene is transcriptional silenced in gastric cancer. Also GKN1 expression at protein level is reduced in patients owing H. pylori-positive chronic gastritis7 and strongly down-regulated or completely absent in patients with gastric cancer8,9. We observed that GKN1 down-regulation is one of the leading causes of gastric cancer (GC) development7,9. In particular, the decrease of GKN1 expression seems to occur throughout the progressive stages of neoplastic transformation and the chronic inflammation of the stomach starts the histopathologic progression of chronic gastritis through gastric atrophy, intestinal metaplasia, dysplasia, and lastly GC9. Over-expression of GKN1 in gastric adenocarcinoma cell lines (AGS and MKN28) activates the expression of Fas receptor, while treatment with an anti-Fas antibody appreciably increases apoptosis10. Moreover, treatment of tumor cells with recombinant human GKN1 (rGKN1) reduceds the proliferation of AGS cells compared with human embryonic kidney cells (HEK 293) and non-gastric cancer cells (H1355)11. These findings denotes that GKN1 is a tumor-suppressor and a modulator of apoptotic signals in GC.
GKN1 and the anti-amyloidogenic property of BRICHOS domain
Beside its role in gastric mucosal protection, GKN1 plays also other important additional functions correlated to the presence in its central region of a conserved BRICHOS domain of near 100 amino acids that has been found also in proteins characterized by several biological functions and disease associations (Fig. 1)12. The term “BRICHOS” is referred to three proteins in which the domain was originally observed: BRI2, expressed in neurons and related to familial British and Danish dementia (FBD and FDD); Chondromodulin-I (ChM-I), a cartilage-specific glycoprotein related to chondrosarcoma and Lung Surfactant Protein C (SP-C), a transmembrane (TM) protein expressed in epithelial type II cells associated to respiratory distress syndrome (RDS)12. Multiple alignment of the amino acid sequence of GKN1 with that of other proteins containing the BRICHOS domain shows very few identities. However, motifs retained to be of functional importance, like D72 and two conservative cysteine residues C95 and C56 involved in disulfide bridges, are present12 and whose function is most likely important for the stability of the protein. The first two conserved cysteines (C95 and C156), are present in all BRICHOS domains, whereas the other two cysteines (C179 and C193), are conserved in all other mammalian gastrokines. In the case of human GKN1, the two latter cysteines are also involved in disulfide bridge as determined by mass spectrometry (MS) performed on rGKN111 (Fig. 2).
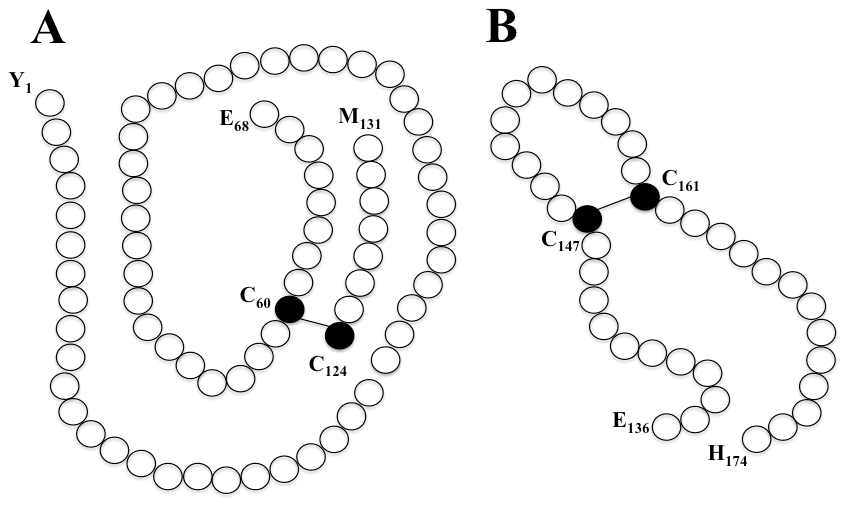
Fig. 2. Formation of disulfide bridges in human recombinant GKN1 (rGKN1).Graphical representation of rGKN1 fragments containing the disulfide bridge in C-terminal region (A) and within the BRICHOS domain (B) as detected by MS after fragmentation of the protein. Amino acid numbering is referred to rGKN111.
In contrast to the low sequence identities, using bio-informatics tools (PSIPRED server v3.3, predict secondary structure), we observed that the BRICHOS domain of GKN1 shows secondary structural features (α-helix and β-sheet) similar to those exhibited by all other proteins containing this domain, like proSP-C and BRI2. In addition, also the 3D model of GKN1 BRICHOS domain constructed at Swiss Model Server11 shows a structural organization approaching that of the corresponding domain of proSP-C13. Both ProSP-C and BRI2 are membrane proteins containing a region with a tendency to form β-sheet structures that can be proteolytically cleaved with mechanisms still unclear. The recombinant BRICHOS domains of these proteins possess a chaperon-like function by precluding fibrils formation of amyloid-beta peptides (1-40) and (1-42)14,15, the products associated to the Alzheimer disease (AD) that are liberated by the amyloid precursor protein (APP)16. The neurophathogenesis of AD is characterized by the accumulation of senile plaques (SPs) and hyper-phosphorylation of tau protein17. SPs are extra cellular deposits composed of various Aβ peptide fragments derived from the amyloidogenic processing of APP that is first cleavage by β-secretase to generate in the extracellular space the APP soluble domain (sAPP) and the membrane-ankored C99 fragment. Subsequently, C99 is processed by the β-secretase at one of several sites varying from +40 to +44 to generate different Aβ peptides among which Aβ(1-40) is the more abundantly form produced both in physiologycal and pathologycal conditions18. The longer peptide Aβ(1-42) is far more prone to oligomerize and forms amyloid fibrils than the shorter peptides19. Although it has been widely accepted that Aβ plays a central role in the onset and progression of AD pathology20,21, accumulating evidence suggests that soluble, non-fibrillar multimeric forms of Aβ peptides (Aβ oligomers, OAβs), rather than insoluble aggregates, might initiate the neuronal dysfunctions associated with the neurodegeneration and cell death22. These toxic oligomeric species are thought to be generated during the amyloidogenic process that involves two nucleation reactions, the primary starting from soluble monomers wherease the secondary nucleation step involves both free monomers and fibrils. As such, the amyloid fibrils are themselves a source for the generation of oligomers that, in turn, can generate new fibrils23. This process could be impaired by the action of specific chaperones that regulate the aggregation mechanism thus preventing amyloid diseases24. Recent studies have shown that BRICHOS domain containig proteins target with highly specificity the secondary nucleation reaction thus it may prevent both the production of toxic oligomers and the new fibrils aggregation25.
Also rGKN1 shows in vitro the property to avoid the aggregation of the peptide Aβ(1-40), as evaluated by Blue Native Page (BN-PAGE)26 (Fig.3) and other methods as described by Altieri et al., 201527. With respect to Aβ(1-40), GKN1 exibits its anti-Aβ aggregation activity at sub-stoichiometric concentrations (1:10 rGKN1/Aβ molar ratio), in accord with data reported for the recombinant proSP-C and BRI2 BRICHOS domains28,29. Regarding the activity of GKN1 toward Aβ(1-42), no data is yet available however, it might be possible that GKN1 behaves similarly to Bri2 and pro-SP-C BRICHOS domains29. It must be pointed out that Aβ(1-42) represents the most AD-relevant amyloid peptide30. The low amount of 1:1 complexes between GKN1 and Aβ(1-40) detected by mass spectrometry indicates that rGKN1 and Aβ(1-40) have a reduced tendency to form a stable complex or perhaps only a small portion of Aβ is accessible for binding to GKN1. The interaction could be mediated by the formation of hydrogen bonds with the –OH group of aromatic amino acids tyrosines that are fully or partially conserved within BRICHOS domains31 and showing structural properties reminiscent of polyphenolic compounds able to retard Aβ aggregation31. Moreover, the affinity constant (KD), evaluated by surface plasma resonance (SPR) technology (3.4x10-5 M), denotes a limited affinity between GKN1 and Aβ and is comparable with that calculated between recombinant proSP-C BRICHOS domain and fibril formation model peptides32. These results might justify i) the low amount of GKN1 (or BRICHOS domain) required to abolish the formation of fibrils and ii) that only a small fraction of the Aβ monomers is present in the GKN1-Aβ complex. The chaperon-like properties of GKN1 could induce conformational change of Aβ that will avoid the formation of toxic species25.
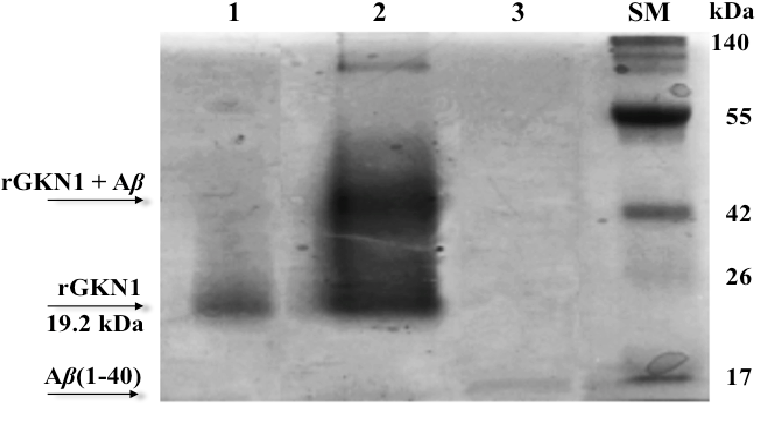
Fig. 3. GKN1 forms a complex with Aβ(1-40).Aβ(1-40) was co-incubated with rGKN1 and their possible aggregation was analyzed under native conditions (native PAGE). A band with a molecular weight greater than that of the individual rGKN1 and Aβ(1-40) was detected after gel staining. Lanes: 1, rGKN1; 2, rGKN1 after incubation with Aβ(1-40); 3, Aβ(1-40) alone. SM, size markers.
GKN1 and its role in APP processing
It has been reported that the product of the dementia gene BRI2 shows the property to inhibit APP processing33,34 and that BRI3, a member of the same gene family, shows the capability to bind APP and to inhibit the production of Aβ35. These BRI protein family, including also BRI1, are transmembrane (TM) proteins of type II. Because also GKN1 is predicted as a type II TM protein (containing the N-terminal inside the cell membrane and 2 transmembrane regions F3-A20 and L158-V174) (TMpred)36, this feature suggested that also GKN1 could interact with APP. In fact, in full-length GKN1 (flGKN1) transfected AGS cells GKN1 was able to bind APP37 and to promote the formation of APP C-terminal Fragments (CTFs) compared with cells treated with a γ-secretase inhibitor (DAPT) or negative control (DMSO). Because CTFs, C83 and C99 are released during APP processing by α- and β-secretases, respectively, these results indicates that the interaction of GKN1 with APP protects APP from γ-secretase. Moreover, pull-down experiments of AGS cell extracts after co-transfection with flGKN1 plus C83 or plus C99 showed that GKN1 directly interacted with CTFs37.
Taking in consideration all the above findings, it may be proposed that GKN1 participates in gastric cells to a twofold mechanism since i) from one side, it displays a chaperone-like activity by delaying amyloid aggregation and ii) from the other side, it binds to the C-terminal region of APP containing the γ-secretase cleavage site, thus inhibiting the processing of CTFs generated by α- and β-secretases activity. In both cases, GKN1 blocks the amyloidogenic pathways.
Beside the important role in gastric mucosal protection1,4,5, the biological reason for the anti-amyloidogenic properties of human GKN1 and for its role in APP processing in gastric tissues is unclear. One explanation might be assigned to the BRICHOS domain acting in general as a chaperon toward peptides with a tendency to asssume β-strand structure. Moreover, the ability of GKN1 to impede γ-secretase cleavadges of APP highlight a possible regulatory role for controlling in normal gastric tissues Aβ formation. However, because the expression of GKN1 is strongly down-regulated or totally absent in GC, the proposed mechanism seems not correlated to gastric carcinogenesis since no amyloidosis was observed by congo red staining in gastric neoplastic tissue (Fig. 4).
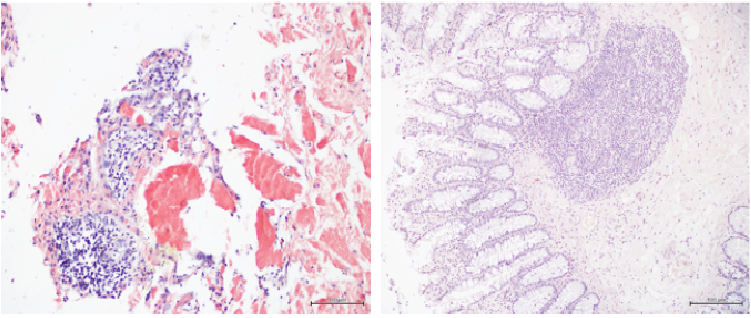
Fig. 4. Red congo staining of human tissues.Left. Congo red deposition of amyloid in AD (nervous tissue). Right. Human gastric tumoral mucosa after congo red staining.
Conclusions
GKN1 is a stomach protein owing the capability to impede the AD Aβ(1-40) peptide aggregation. This property appears attribuable to GKN1 BRICHOS domain that exert a chaperon-like activity for protecting and avoiding peptide aggregation or for transport or secretion of amyloid fibrils27,37. Also the capability of GKN1 to bind APP could play a regulatory role in the processing of APP since it blocks the γ-secretases sites and thus enhancing the non-amyloidogenic pathway37. The overexpression of GKN1 BRICHOS domain in affected tissues might suppress the generation of toxis species. Therefore, considering that the major developments in AD treatment are the amyloid and tau based therapeutics30,38, BRICHOS domain of GKN1 could represents a promising pharmacological tool against amyloid diseases.
Acknowledgments
This work was supported by funds from Programmi di Ricerca Scientifica di Rilevante Interesse Nazionale (2008BKRFBH_003), from FIRB N_RBNE08NKH7_003 and from PON Ricerca e Competitività 2007-2013 (PON01_02782).
Conflicts of Interest
The authors declare no conflict of interest.
Authors’ Contributions
CSDS, FA, GMin and GMis were involved in all experiments, ER was involved in the preparation of a draft manuscript, PA take care of reading and approval of the final manuscript.
References
- Toback FG, Walsh Reitz MM, Musch MW, et al. Peptide fragments of AMP-18 a novel secreted gastric antrum mucosal protein are mitogenic and motogenic. Am J Physiol Gastrointest Liver Physiol. 2003; 285: G344–G353.
- Martin TE, Powell CT, Wang Z, et al. A novel mitogenic protein that is highly expressed in cells of the gastric antrum mucosa. AJP Gastrointesinal and Liver Physiology. 2003; G332-G343.
- Shiozaki K, Nakamori S, Tsujie M, et al. Human stomach-specific gene CA11 is downregulated in gastric cancer. Int J Oncol. 2001; 701-707.
- Walsh-Reitz MM, Huang EF, Musch MW, et al. AMP-18 protects barrier function of colonic epithelial cells: Role of tight junction proteins. Am J Physiol Gastroint Liver Physiol. 2005; 289: G163-G171.
- Rippa E, Altieri F, Di Stadio CS, et al. Ectopic expression of gastrokine 1 in gastric cancer cells up-regulates tight and adherens junction proteins network. Pathol Res Pract. 2015; 211(8): 577-83.
- Oien KA, McGregor F, Butler S, et al. Gastrokine 1 is abundantly and specifically expressed in superficial gastric epithelium, down-regulated in gastric carcinoma, and shows high evolutionary conservation. J Pathol. 2004; 203: 789-797.
- Rippa E, Martin G, Rocco A, et al. Changes of protein expression in Helicobacter pylori-infected human gastric mucosa. Current Topics in Peptide & Protein Research. 2007; 8: 35-43.
- Nardone G, Rippa E, Martin G, et al. Gastrokine 1 expression in patients with and without Helicobacter pylori infection. Digestive and Liver Disease. 2007; 39: 122-129.
- Nardone, G, Martin, G, Rocco, A, et al. Molecular expression of gastrokine 1 in normal mucosa and in Helicobacter pylori related preneoplastic and neoplastic gastric lesions. Cancer Biology and Therapy. 2008; 7: 1890-1895.
- Rippa E, La Monica G, Allocca R, et al. Overexpression of gastrokine 1 in gastric cancer cells induces Fas-mediated apoptosis. J Cell Physiol. 2011; 226: 2571–2578.
- Pavone LM, Del Vecchio P, Mallardo P, et al. Structural characterization and biological properties of human gastrokine 1. Mol Biosyst. 2013; 9: 412-421.
- Sanchez Pulido L, Devos D, Valencia A. BRICHOS: a conserved domain in proteins associated with dementia, respiratory distress and cancer. Trends biochem Sci. 2002; 329-332.
- Willander H, Askarieh G, Landreh M, et al. High-resolution structure of a BRICHOS domain and ist implications for antiamyloid chaperone activity on lung surfactant protein C. Proc Natl Acad Sci. 2012; 109(7): 2325-2329.
- Johansson H, Nordling K, Weaver TE, et al. The Brichos domain containing C-terminal part of pro-surfactant protein C binds to an unfolded poly-val transmembrane segment. J Biol Chem. 2006; 281: 21032-21039.
- Peng S, Fitzen M, Jornvall H, et al. The extracellular domain of Bri2 (ITM2B) binds the ABri peptide (1-23) and amyloid b-peptide (Ab1-40): Implications for Bri2 effects on processing of amyloid precursor protein and Ab aggregation. Biochem Biophys Res Commun. 2010; 393: 356-361.
- Willander H, Hermansson E, Johansson J, et al. BRICHOS domain associated with lung fibrosis, dementia and cancer e a chaperone that prevents amyloid fibril formation. FEBS J. 2011; 278 (20): 3893-3904.
- Ezoulin MJ, Ombetta JE, Dutertre-Catella H, et al. Antioxidative properties of galantamine on neuronal damage induced by hydrogen peroxide in SK–N–SH cells. Neurotoxicology. 2008; 29: 270–277.
- Haass C, Kaether C, Thinakaran G, et al. Trafficking and proteolytic processing of APP. Cold Spring Harb Perspect Med. 2012; 2(5): a006270.
- Thinakaran G, Koo EH. Amyloid precursor protein trafficking processing and function. J Biol Chem. 2008; 283(44): 29615-296159.
- Hardy JA, Higgins GA. Alzheimer's disease: the amyloid cascade hypothesis. Science. 1992; 256(5054): 184-185.
- Selkoe DJ. Alzheimer's disease: a central role for amyloid. J Neuropathol Exp Neurol. 1994; 53(5): 438-447.
- Benilova I, Karran E, De Strooper B. The toxic Aβ oligomer and Alzheimer's disease: an emperor in need of clothes. Nat Neurosci. 2012; 15(3): 349-357.
- Cohen SI, Linse S, Luheshi LM, et al. Proliferation of amyloid-β42 aggregates occurs through a secondary nucleation mechanism. Proc Natl Acad Sci. 2013; 110: 9758–9763.
- Landreh M, Rising A, Presto J, et al. Specific chaperones and regulatory domains in control of amyloid formation. J Biol Chem. 2015; 290(44): 26430-26436.
- Cohen SI, Arosio P, Presto J, et al. A molecular chaperone breaks the catalytic cycle that generates toxic Aβ oligomers. Nat Struct Mol Biol. 2015; 22: 207–213.
- Swamy M, Siegers GM, Minguet S, et al. Blue native polyacrylamide gel electrophoresis (BN-PAGE) for the identification and analysis of multiprotein complexes. Sci STKE. 2006; 345: pl4.
- Altieri F, Di Stadio CS, Severino V, et al. Anti-amyloidogenic property of human gastrokine 1. Biochimie. 2014; 106: 91-100.
- Nerelius C, Gustafsson M, Nordling K, et al. Anti-amyloid activity of the c-terminal domain of proSP-c against amyloid b-peptide and Medin. Biochemistry. 2009; 48: 3778-3786.
- Willander H, Presto J, Askarieh G, et al. BRICHOS domains efficiently delay fibrillation of amyloid b-peptide. J Biol Chem. 2012; 287(37): 31608-31617.
- Willian O, Castillo, Andrés F, et al. Galanthamine decreases genotoxicity and cell death induced by b-amyloid peptide in SH-SY5Y cell line. NeuroToxicology. 2016; 57: 291–297.
- Knight SD, Presto J, Linse S, et al. The BRICHOS domain, amyloid fibril formation, and heir relationship. Biochemistry. 2013; 53: 7523–7531.
- Fitzen M, Alvelius G, Nordling K, et al. Peptide-binding specificity of the prosurfactant protein C Brichos domain analyzed by electrospray ionization mass spectrometry. Rapid Commun Mass Spectrom. 2009; 23(22): 3591-3598.
- Fotinopoulou A, Tsachaki M, Vlavaki M, et al. BRI2 interacts with amyloid precursor protein (APP) and regulates amyloid b (Ab) production. J Biol Chem. 2005; 280(35): 30768-30772.
- Tsachaki M, Fotinopoulou A, Slavi N, et al. BRI2 interacts with BACE1 and regulates its cellular levels by promoting its degradation and reducing its mRNA levels. Curr Alzheimer Res. 2013; 10(5): 532-541.
- Matsuda S, Matsuda Y, D'Adamio L. BRI3 inhibits amyloid precursor protein processing in a mechanistically distinct manner from its homologue dementia gene BRI2. J Biol Chem. 2009; 284(23): 15815-15825.
- K. Hofmann & W. Stoffel. TMbase - A database of membrane spanning proteins segments Biol. Chem. Hoppe-Seyler. 1993; 374:166.
- Di Stadio CS, Altieri F, Minopoli G, et al. Role of human GKN1 on APP processing in gastric cancer. Biochimie. 2017; 135: 149-153.
- Anand R, Gill KD, Mahdi AA. Therapeutics of Alzheimer’s disease Past present and future. Neuropharmacology. 2014; 7627-7650.